Latin Name
Common Name
Found 66 Resources
Page 1 of 1
Page 1 of 1



Asian Spongy Moth
Lymantria dispar
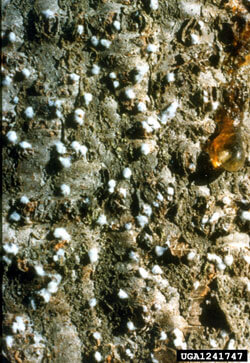









Cactus Moth
Cactoblastis cactorum










European Larch Canker
Lachnellula (Dasyscypha)
European Oak Bark Beetle
Scolytus intricatus
European Spongy Moth
Lymantria dispar

Fusarium euwallacea
Fusarium euwallacea

Harrisia Cactus Mealybug
Hypogeococcus pungens

Kuroshio Shot Hole Borer
Euwallacea kuroshio





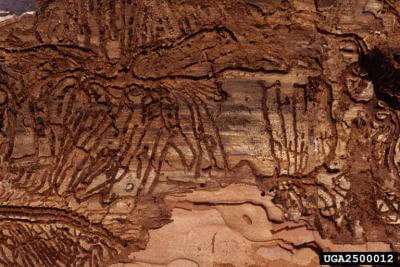
Oak Dieback
Phytophthora quercina


Oystershell scale
Lepidosaphes ulmi



Pine Flat Bug
Aradus cinnamomeus